
ヒト肝キメラマウスの特性
 日本人科学者の多大な貢献によって、ヒト肝キメラマウスが開発されました。作成方法の原理と手順は、弊社ホームページの「続・肝炎ウイルス十話」に掲載されている第六話「キメラマウスモデルのヒト肝炎ウイルス感染」をご覧ください。キメラマウスの代名詞として「肝臓をヒト肝細胞に置き換えたマウス」があります。しかし肝臓は肝細胞だけで構成されている訳ではないのです(図1)。流入血管が二系統(肝動脈と門脈)流出血管が一系統(中心静脈)あり、胆汁が流出する胆細管と共に複雑に設計された「肝小葉」が単位となって、コンパクトに組み立てられています。内皮下の間隙(ディッセ腔とも呼ばれています)には、日本の医学者が発見した星(伊東)細胞があり、肝硬変の原因となる線維化を担当しています。これら全てがマウス由来で、ヒト肝細胞が借用しています。
日本人科学者の多大な貢献によって、ヒト肝キメラマウスが開発されました。作成方法の原理と手順は、弊社ホームページの「続・肝炎ウイルス十話」に掲載されている第六話「キメラマウスモデルのヒト肝炎ウイルス感染」をご覧ください。キメラマウスの代名詞として「肝臓をヒト肝細胞に置き換えたマウス」があります。しかし肝臓は肝細胞だけで構成されている訳ではないのです(図1)。流入血管が二系統(肝動脈と門脈)流出血管が一系統(中心静脈)あり、胆汁が流出する胆細管と共に複雑に設計された「肝小葉」が単位となって、コンパクトに組み立てられています。内皮下の間隙(ディッセ腔とも呼ばれています)には、日本の医学者が発見した星(伊東)細胞があり、肝硬変の原因となる線維化を担当しています。これら全てがマウス由来で、ヒト肝細胞が借用しています。
移植された100万(106)個のヒト肝細胞は2ヶ月の間に7〜10回分裂(倍増)を繰り返し約100倍にもなって、病変のある白い自己肝臓を置換します。キメラマウス開発当初の予想は1回の移植で100倍増する「ねずみ算」的な増殖で、ひとたび成功すれば移植源としての肝細胞が永代確保される見込みでした。しかし継代移植を繰り返すうちに血清中ヒトアルブミンから算出される、マウス肝のヒト肝細胞・置換率が、次第に低下することが判明しました。
細胞の分裂には遺伝子を構成するDNAの複製が必要です。しかしDNAの先端にあり複製を開始する塩基配列(テロメアと呼ばれています)が、あたかも「命の回数券」のように、一回の分裂で1単位ずつ切り離されます。回数券は50枚しかないので、肝細胞が未来永劫に分裂を繰り返す訳にはいかないらしいのです。恐らくこれが原因となって、ドナーの年齢がキメラマウスのヒト肝細胞置換率を決定するのでしょう。高い置換率(70%以上)が期待できるのはドナー年齢がせいぜいローティーン迄で、30歳以上ではとても望めないようです。
ウイルス性肝疾患の進展速度は種々の宿主因子によって支配され、その一つとして人種があります。特に黒人のC型慢性肝炎患者にはインターフェロン療法が効きにくいことが知られています。カタログ販売されている肝細胞は1,000万(107)個が1単位となっていますので、一人のドナーから約500単位が得られ、1単位でキメラマウス10匹分ありますから、全部で5,000匹に相当します。黒色人種(ネグロイド)、白色人種(コーカソイド)と黄色人種(モンゴロイド)の全部が揃っていますので、人種別のC型慢性肝炎・薬剤抵抗性を解明する手がかりが得られるかもしれません。
キメラマウス作成の鍵は、ヒト肝細胞の再生を促進するために自己(マウス)肝細胞を破壊する仕組みが施されていることです。肝障害があるので、移植されたヒト肝細胞は際限なく増殖を続けて体重15グラムで1.5グラム(10%)以上を占め、ヒトで約2%(体重60キロで1.5キロ)までの自律的成長とはかけ離れた世界です。勿論、移植を成立させるための免疫欠如があります。しかも、図1のように、肝細胞以外は、全てがマウスからの借り物です。従って、キメラマウスが最も得意とする独壇場は肝細胞だけを対象とする局面です。それには少なくとも三通りがあります。(1)肝炎ウイルスのヒト肝細胞に対する感染性の判定とその予防法の確立、(2)抗ウイルス薬剤の効果判定、と(3)あらゆる新薬開発に必要な薬剤副作用の判定です。今回は、それぞれを順番にお話ししたいと思います。
ヒト肝炎ウイルスによる肝細胞の感染
 血液を介して感染し、慢性肝疾患を起こす肝炎ウイルス(B型肝炎ウイルス [HBV] とC型肝炎ウイルス [HCV])には厳密な種特異性があるので、ヒト以外には系統発生学的に近縁のチンパンジーにしか伝搬しません。しかしチンパンジーはもはや絶滅危惧種となり、倫理的配慮もあって日本での感染実験は平成18年度末(2007年3月)に打ち切られました。これを待っていたかのように颯爽と登場したのがキメラマウスです。日本の肝細胞癌では、90%以上でウイルス性肝炎が原因となります。ウイルス性肝疾患は所詮HBVとHCVがもたらす感染症ですので、感染性の評価には肝細胞だけあれば、こと足ります。体内の全組織で、肝炎ウイルスに感染する唯一の細胞だからです。これまで感染性を判定する「リトマス試験紙」の役割は、ひとえにチンパンジーに任されてきました。HBVの研究に数百頭ものチンパンジーが使われ、HCV発見にもそれ以上が供されてきました。体重15グラムの可憐なキメラマウスが、一対一の関係で、体重60キログラムを超え飼育費用だけで年間五百万円はかかる、チンパンジーの代役を立派に務めると、誰が予測できたでしょうか。
血液を介して感染し、慢性肝疾患を起こす肝炎ウイルス(B型肝炎ウイルス [HBV] とC型肝炎ウイルス [HCV])には厳密な種特異性があるので、ヒト以外には系統発生学的に近縁のチンパンジーにしか伝搬しません。しかしチンパンジーはもはや絶滅危惧種となり、倫理的配慮もあって日本での感染実験は平成18年度末(2007年3月)に打ち切られました。これを待っていたかのように颯爽と登場したのがキメラマウスです。日本の肝細胞癌では、90%以上でウイルス性肝炎が原因となります。ウイルス性肝疾患は所詮HBVとHCVがもたらす感染症ですので、感染性の評価には肝細胞だけあれば、こと足ります。体内の全組織で、肝炎ウイルスに感染する唯一の細胞だからです。これまで感染性を判定する「リトマス試験紙」の役割は、ひとえにチンパンジーに任されてきました。HBVの研究に数百頭ものチンパンジーが使われ、HCV発見にもそれ以上が供されてきました。体重15グラムの可憐なキメラマウスが、一対一の関係で、体重60キログラムを超え飼育費用だけで年間五百万円はかかる、チンパンジーの代役を立派に務めると、誰が予測できたでしょうか。
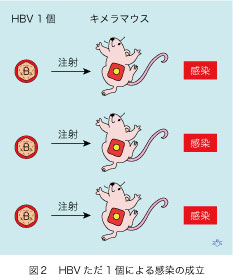
肝炎ウイルスがただ一個あれば、肝細胞一個が先ず感染し、そこでウイルスが増殖して持続感染が成立する、と理論的には考えられます。しかし、チンパンジーでの感染実験では一頭を感染させるために、少なくとも十個のHBV(HCVも同様)が必要でした。この問題は、キメラマウスで解決しました(図2)。三匹のキメラマウスのそれぞれに、唯一個のHBVを注射してから4週後に、全てが感染して以後、血中にHBV DNAが高い濃度で持続しました。
HBVゲノタイプが増殖速度に及ぼす影響
 B型肝炎ウイルス(HBV)にはアルファベット大文字のAからH迄で標記される、8種類の遺伝子型(ゲノタイプ)があり、約3,200個の塩基配列が互いに8%以上(256個に相当します)違っていることから識別されています。中でもゲノタイプAには、際立った特徴があります。成人になってからのHBV感染は一過性に終わり、持続することは稀です。ところがA型に限っては、10%以上で感染が持続するのです。近年日本でも不特定多数との異性・同性間の性的接触の結果としてゲノタイプA型のHBV感染が増加しています。初回献血者に発見されるHBVもゲノタイプAの割合が高く、特に大都会とその周辺でその傾向が高いのです(50%以上にも及ぶ所があります)。ゲノタイプA型には、いくつかの亜型がありますが、日本では欧米型のAe(A2とも呼ばれます)が殆どです。
B型肝炎ウイルス(HBV)にはアルファベット大文字のAからH迄で標記される、8種類の遺伝子型(ゲノタイプ)があり、約3,200個の塩基配列が互いに8%以上(256個に相当します)違っていることから識別されています。中でもゲノタイプAには、際立った特徴があります。成人になってからのHBV感染は一過性に終わり、持続することは稀です。ところがA型に限っては、10%以上で感染が持続するのです。近年日本でも不特定多数との異性・同性間の性的接触の結果としてゲノタイプA型のHBV感染が増加しています。初回献血者に発見されるHBVもゲノタイプAの割合が高く、特に大都会とその周辺でその傾向が高いのです(50%以上にも及ぶ所があります)。ゲノタイプA型には、いくつかの亜型がありますが、日本では欧米型のAe(A2とも呼ばれます)が殆どです。
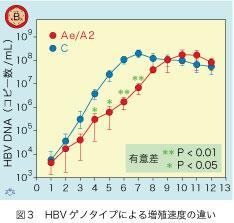
ゲノタイプAのHBV初感染が何故持続するかが、キメラマウスでの感染実験から追求されました。キメラマウス3匹ずつに105個のAe型と、日本で一番頻度の高いゲノタイプC型のHBVを静脈注射して、以後毎週血液中のHBV DNAが追跡されました(図3)。時間が経つにつれて、Ae型HBVの増殖速度がC型と比べて有意に低下しています。対数的(exponential)な増殖をする期間で直線を引き、その勾配からHBVが10倍に増殖するのに必要な日数(log time)を知ることができます。Ae型は12.7日、C型は7.8日と算出されますので、増殖速度に約2倍の開きがあります。この結果から、Ae型が遅い初期増殖速度を介して、成人HBV初感染後の感染持続を助長していることが理解できます。Ae型は増殖のスピードが遅いので、宿主の免疫排除機構が低下し、そのために成人感染でもHBV感染が持続することが多いのでしょう。
HCV感染防御抗体の開発と実用性
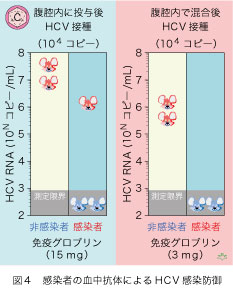 肝炎ウイルス対策の最良の道は、感染を防止することです。感染を防御できる中和抗体の存在が、26年以前にHCVに感染した患者で証明されました(図4)。ただし、感染当初のHCV株に対する中和抗体である、という制約があります。この患者は今でもHCVに持続感染していますから、現在感染中のHCVに対する中和抗体は持っていないのです。単一のHCV株(ゲノタイプ1a型)であることに加えて、感染の予防に必要な高い抗体量には実現性がなく、しかもHCVは腹腔内接種なのでかなり「荒っぽい仕事」のようです。その上、最良の「お膳立て」であったのに、3匹中の1匹が感染してしまいました。抗体とHCVを腹腔内に同時投与した2匹は、ともに感染を免れましたが、臨床応用にはほど遠い設定です。
肝炎ウイルス対策の最良の道は、感染を防止することです。感染を防御できる中和抗体の存在が、26年以前にHCVに感染した患者で証明されました(図4)。ただし、感染当初のHCV株に対する中和抗体である、という制約があります。この患者は今でもHCVに持続感染していますから、現在感染中のHCVに対する中和抗体は持っていないのです。単一のHCV株(ゲノタイプ1a型)であることに加えて、感染の予防に必要な高い抗体量には実現性がなく、しかもHCVは腹腔内接種なのでかなり「荒っぽい仕事」のようです。その上、最良の「お膳立て」であったのに、3匹中の1匹が感染してしまいました。抗体とHCVを腹腔内に同時投与した2匹は、ともに感染を免れましたが、臨床応用にはほど遠い設定です。
 HCV持続感染の究極の治療法として、肝移植があります。最大の難関として、移植した肝臓がHCVに100%再感染する現実があります。HBV持続感染では、HBs抗体(抗・表面抗原抗体)の術前、術中と術後投与が、完璧に近く再感染を防御しますので、HCVの再感染も抗体で阻止できる可能性が残されています。前提は、HCVが肝細胞の表面にある受容体と結合する部位(HCVのE2タンパク)を遮断できる抗体を無制限に調達して、投与法ならびに感染予防効果を実証することです。このためには量産が可能なモノクローナル抗体で、HCVのゲノタイプと多様性(quasispecies)に関係なく、あらゆる種類のHCVを中和する抗体が求められます。予防効果の最終判定には、キメラマウスをおいて他ありません。
HCV持続感染の究極の治療法として、肝移植があります。最大の難関として、移植した肝臓がHCVに100%再感染する現実があります。HBV持続感染では、HBs抗体(抗・表面抗原抗体)の術前、術中と術後投与が、完璧に近く再感染を防御しますので、HCVの再感染も抗体で阻止できる可能性が残されています。前提は、HCVが肝細胞の表面にある受容体と結合する部位(HCVのE2タンパク)を遮断できる抗体を無制限に調達して、投与法ならびに感染予防効果を実証することです。このためには量産が可能なモノクローナル抗体で、HCVのゲノタイプと多様性(quasispecies)に関係なく、あらゆる種類のHCVを中和する抗体が求められます。予防効果の最終判定には、キメラマウスをおいて他ありません。
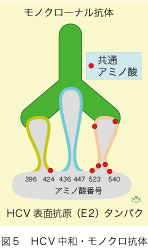
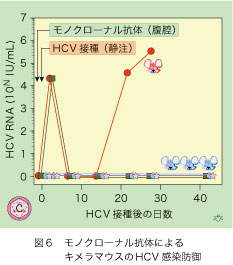 HCV感染者の骨髄に由来した個々の免疫細胞からE2タンパクと結合するモノクローナル抗体(唯一つの抗原基を識別する均一な抗体)が得られ、これはE2タンパクの三つの離れた部位が構成する構造抗原基(conformational epitope)を認識し、 全てのHCV株に共通したアミノ酸を、多数含んでいました(図5)。このモノクローナル抗体(3mg)を腹腔内に投与した翌日にHCV(ゲノタイプ1a型、 105コピー)を静注した4匹のキメラマウスで、血液中のHCV RNAが追跡されました(図6)。数日後に2匹が一過性感染し、そのうち1匹では20日後に突破感染が起こって、接種後30日に衰弱死しました。
HCV感染者の骨髄に由来した個々の免疫細胞からE2タンパクと結合するモノクローナル抗体(唯一つの抗原基を識別する均一な抗体)が得られ、これはE2タンパクの三つの離れた部位が構成する構造抗原基(conformational epitope)を認識し、 全てのHCV株に共通したアミノ酸を、多数含んでいました(図5)。このモノクローナル抗体(3mg)を腹腔内に投与した翌日にHCV(ゲノタイプ1a型、 105コピー)を静注した4匹のキメラマウスで、血液中のHCV RNAが追跡されました(図6)。数日後に2匹が一過性感染し、そのうち1匹では20日後に突破感染が起こって、接種後30日に衰弱死しました。
でも、一過性感染した1匹を含む残りの3匹は、40日を超える観察期間中にわたってHCV感染を免れたのです(従って、防御率は75%となります)。将来この方向の研究には希望がもてそうで、ヒト肝キメラマウスの活躍が期待できます。
HBVの新種ラミブジン耐性・変異体
 ラミブジンは核酸同族体の一種でHBVの複製を強力に阻害し、副作用が少ないのでB型慢性肝炎の長期治療に使われています。しかし、ラミブジンに耐性がある変異体(ミュータント)が1年で24%、4年では70%もの高頻度で出現する難点があります。殆どの変異体は、ポリメラーゼの特定部位(YMDDモチーフ)のアミノ酸置換を持っています。ラミブジン治療中にHBV DNAが上昇し肝炎が増悪した1症例では塩基配列を調べても、YMDDモチーフに変異が見られませんでした。ところが他の部位に変異があり、ポリメラーゼ第181番目のアミノ酸が、アラニンからスレオニンに置換されていました。しかし、それが原因であることを実証する手だてがなかったのです。
ラミブジンは核酸同族体の一種でHBVの複製を強力に阻害し、副作用が少ないのでB型慢性肝炎の長期治療に使われています。しかし、ラミブジンに耐性がある変異体(ミュータント)が1年で24%、4年では70%もの高頻度で出現する難点があります。殆どの変異体は、ポリメラーゼの特定部位(YMDDモチーフ)のアミノ酸置換を持っています。ラミブジン治療中にHBV DNAが上昇し肝炎が増悪した1症例では塩基配列を調べても、YMDDモチーフに変異が見られませんでした。ところが他の部位に変異があり、ポリメラーゼ第181番目のアミノ酸が、アラニンからスレオニンに置換されていました。しかし、それが原因であることを実証する手だてがなかったのです。
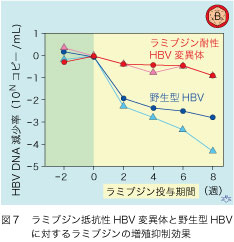
キメラマウスはこのような局面で最大の力を発揮します。このアミノ酸変異のあるラミブジン抵抗性のHBV変異体と変異のない野生型HBVが、2匹ずつのキメラマウスに接種されました(図7)。毎日餌の中に キログラム体重あたり30ミリグラムを混入させましたが、マウスはラミブジンが嫌いなようで、食べさせるのに苦労したようです。患者での治療常用量は1日100ミリグラムなので、体重比で10倍は多く飲まされたことになります。ラミブジン抵抗性のHBV変異体を接種されたマウスでは血中HBV DNA値に殆ど変化がなかったのですが、野生型HBV接種マウスでは大幅な低下が見られ、アミノ酸変異と薬剤抵抗性との因果関係が、見事に実証されました。
HCVゲノタイプが治療応答に及ぼす影響
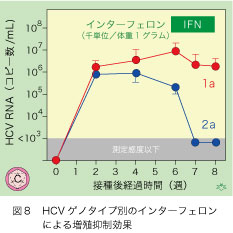 C型肝炎ウイルスには1型から6型までのゲノタイプがあり日本人C型慢性肝炎患者には1型が70%、残りは2型が感染しています。インターフェロン・リバビリン併用療法の治療応答は大きく違っていて、ゲノタイプ1型では50%に止まるのに対して、2型では80%以上に及びます。応答は、治療が終了してから6ヶ月以上にわたって血中のHCV RNA陰性が持続することで判定され、これがあれば99%以上の確率で応答が4年間以上も持続します。1a型および2a型のHCVクローンに感染したキメラマウス血清を接種された各3匹の間には、HCV RNAの減少に大きな違いが認められました(図8)。人体でゲノタイプ1型が2型に比べて治療応答が悪いことが、実験的にも実証されて、「納得!」が得られました。
C型肝炎ウイルスには1型から6型までのゲノタイプがあり日本人C型慢性肝炎患者には1型が70%、残りは2型が感染しています。インターフェロン・リバビリン併用療法の治療応答は大きく違っていて、ゲノタイプ1型では50%に止まるのに対して、2型では80%以上に及びます。応答は、治療が終了してから6ヶ月以上にわたって血中のHCV RNA陰性が持続することで判定され、これがあれば99%以上の確率で応答が4年間以上も持続します。1a型および2a型のHCVクローンに感染したキメラマウス血清を接種された各3匹の間には、HCV RNAの減少に大きな違いが認められました(図8)。人体でゲノタイプ1型が2型に比べて治療応答が悪いことが、実験的にも実証されて、「納得!」が得られました。
ウイルス性慢性肝炎・新治療薬開発への応用
C型慢性肝炎の薬物療法は、近年大きく進歩しました。しかし難治性であるゲノタイプ1型HCV感染患者では、依然として約半数でしかウイルス排除効果がないので、とても満足すべき状態ではありません。従って、新たな治療薬の登場が待ち望まれています。抗・肝炎ウイルス剤の開発は、正しくキメラマウスの独壇場です。先ずは置き換わったヒト肝細胞に感染している肝炎ウイルスの増殖を抑制する効果を、試験管内(in vitro)ではなく、生体内(in vivo)で判定することができます。それに加えて、新薬開発上で大きな障壁となる、薬剤副作用の有無と程度をも、調べることが可能なのです。
全ての薬剤は肝臓で代謝され、必要となれば解毒されます。キメラマウスの血液検査から、ヒト肝細胞が産生するほぼ全てのタンパクが検出されています。とりわけ薬剤代謝の中心となるチトクローム酵素系は、全てヒト由来であることが、薬剤の副作用判定を容易で正確にしています。実は、これがヒト肝キメラマウスの最大の「売り」なのです。従来のマウスとラットを使った検査法では、チトクローム酵素の種差のために、ヒトで薬害があるか否かを正しく判定することができませんでした。今のところ日本では主として肝炎ウイルスの研究に使われていますが、肝臓関係に限らず、あらゆる新薬の副作用を試験的治療(治験)開始前に判定することができます。新薬開発が、今後キメラマウス最大の応用になることが予想できます。
「背が高くなければ、スーパーモデルになれない」とは云わせない
 文字通り、キメラマウスは今や小さなスーパー(実験動物)モデルの大スターとなりました。でも、スーパーモデルには共通した悩みがあります。完璧な容姿は、健康の維持と高い知的活動を必ずしも保証しません。キメラマウスは肝炎ウイルス(特にHCV)感染モデルとして、他の追随を許しません。しかし生存期間が短く(感染後数ヶ月)免疫反応が欠如しているので、ウイルス性肝炎と肝細胞癌の発生病理を解明することは望めません。一方で、あらゆる局面がスーパーモデルを求めている訳ではありません。ニューヨーク・ロンドン・ミラノ・パリ・東京の五大コレクションだけが人類のファッションではないのです。任せられる所は適材適所の一般モデルに譲って、たとえシャッチョコダチしても真似できない場面だけに、颯爽と登場するのが「真打ち」の醍醐味ではないでしょうか。
文字通り、キメラマウスは今や小さなスーパー(実験動物)モデルの大スターとなりました。でも、スーパーモデルには共通した悩みがあります。完璧な容姿は、健康の維持と高い知的活動を必ずしも保証しません。キメラマウスは肝炎ウイルス(特にHCV)感染モデルとして、他の追随を許しません。しかし生存期間が短く(感染後数ヶ月)免疫反応が欠如しているので、ウイルス性肝炎と肝細胞癌の発生病理を解明することは望めません。一方で、あらゆる局面がスーパーモデルを求めている訳ではありません。ニューヨーク・ロンドン・ミラノ・パリ・東京の五大コレクションだけが人類のファッションではないのです。任せられる所は適材適所の一般モデルに譲って、たとえシャッチョコダチしても真似できない場面だけに、颯爽と登場するのが「真打ち」の醍醐味ではないでしょうか。
参考文献
Tabuchi A, Tanaka J, Katayama K, Mizui M, Matsukura H, Yugi H, Shimada T, Miyakawa Y, Yoshizawa H: Titration of hepatitis B virus infectivity in the sera of pre-acute and late acute phases of HBV infection: transmission experiments to chimeric mice with human liver repopulated hepatocytes. J Med Virol 2008;80:2064-2068.
Sugiyama M, Tanaka Y, Kato T, Orito E, Ito K, Acharya SK, Gish RG, Kramvis A, Shimada T, Izumi N, Kaito M, Miyakawa Y, Mizokami M: Influence of hepatitis B virus genotypes on the intra- and extracellular expression of viral DNA and antigens. Hepatology 2006;44:915-924.
Vanwolleghem T, Bukh J, Meuleman P, Desombere I, Meunier JC, Alter H, Purcell RH, Leroux-Roels G: Polyclonal immunoglobulins from a chronic hepatitis C virus patient protect human liver-chimeric mice from infection with a homologous hepatitis C virus strain. Hepatology 2008;47:1846-1855.
Law M, Maruyama T, Lewis J, Giang E, Tarr AW, Stamataki Z, Gastaminza P, Chisari FV, Jones IM, Fox RI, Ball JK, McKeating JA, Kneteman NM, Burton DR: Broadly neutralizing antibodies protect against hepatitis C virus quasispecies challenge. Nat Med 2008;14:25-27.
Yatsuji H, Noguchi C, Hiraga N, Mori N, Tsuge M, Imamura M, Takahashi S, Iwao E, Fujimoto Y, Ochi H, Abe H, Maekawa T, Tateno C, Yoshizato K, Suzuki F, Kumada H, Chayama K: Emergence of a novel lamivudine-resistant hepatitis B virus variant with a substitution outside the YMDD motif. Antimicrob Agents Chemother 2006;50:3867-3874.
Hiraga N, Imamura M, Tsuge M, Noguchi C, Takahashi S, Iwao E, Fujimoto Y, Abe H, Maekawa T, Ochi H, Tateno C, Yoshizato K, Sakai A, Sakai Y, Honda M, Kaneko S, Wakita T, Chayama K: Infection of human hepatocyte chimeric mouse with genetically engineered hepatitis C virus and its susceptibility to interferon. FEBS Lett 2007;581:1983-1987.
![]()